高等植物の形態形成における膜交通メカニズムの役割
真核生物の形態形成には,細胞の極性確立が非常に重要であることが知られています。細胞極性の確立には,細胞膜上へのタンパク質の方向性を持った輸送,いわゆる極性輸送が重要な働きをしています。植物におけるタンパク質の極性輸送は,根毛や花粉管の伸長,いわゆる“先端成長”をモデルとして精力的に研究され,カルシウム,細胞骨格,ミオシン等モータータンパク質,フォスファチジルイノシトールリン脂質,Rab, Ropなどの低分子量GTPase,SNAREなどの分子が重要な働きをしていることがわかっていますが,これらの分子間の相互作用に関しては,不明な点が多い状況です。
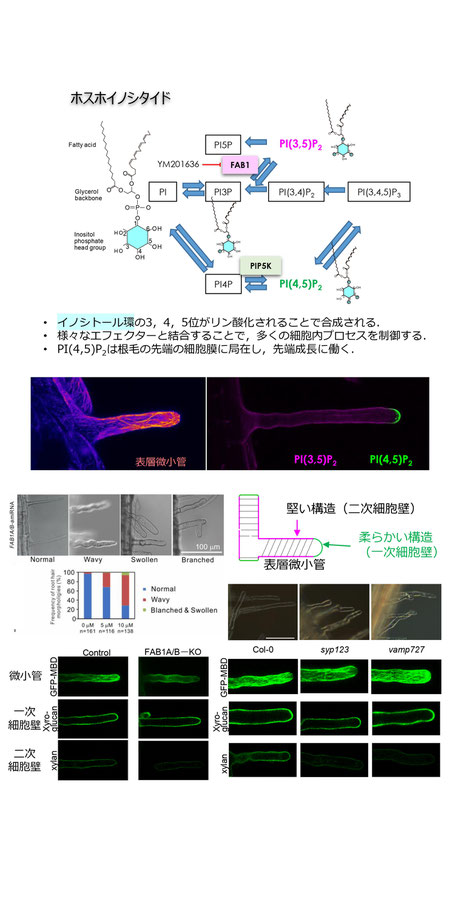
当研究室では,1994年に、シロイヌナズナから、液胞膜SNARE,AtVam3の単離および解析を世界に先駆けて行った研究を皮切りに,液胞の動的構造の解析,シロイヌナズナSNARE細胞内局在マップの作成,細胞膜SNAREの発現および細胞膜上での局在解析,根毛と花粉管伸長に関与するSANREの発見,フォスファチジルイノシトール3リン酸(PI3P)をフォスファチジルイノシトール3,5ビスリン酸に変換するキナーゼFAB1とエンドサイトーシスとの関わりなどの成果を得てきました。
特に,我々は,根毛の伸長過程における根毛特異的SNARE, SYP123および全細胞で普遍的に発現しているSYP132についての機能解析を進め,SYP123は,アクチン繊維依存的に根毛の先端に集中し,輸送小胞に存在するR-SNAREの一種,AtVAMP721とSNARE複合体を形成し,一方,SYP132は,細胞膜全体に均一に存在し,やはりAtVAMP721とSNARE複合体を形成して,BFA非感受性の新規タンパク質のエキソサイトーシスに関与していることを明らかにしました。
一方,イノシトールリン脂質(PIs)は,イノシトール環の3,4,5位の水酸基がリン酸結合されることにより,7種類存在します.そのうち,3位と5位にリン酸が結合したPI(3,5)P2は,FAB1によって合成され,4位と5位にリン酸が結合したPI(4,5)P2は,PIP5Kによって合成されることが知られています.
我々は,モデル植物シロイヌナズナを用いて,FAB1の機能解析を中心に研究を行い,FAB1が後期エンドソームに局在し,後期エンドソームの成熟に働き,加えて,表層微小管の配向制御に機能することを明らかにしてきました.